





Introduction
There are several sources of reactive oxygen in the human body. Production of superoxide in mitochondria is a by-product of the function of the respiratory chain [ 1 ]. The first known example of regulated generation of reactive oxygen species (ROS) in mammalian cells was through the respiratory burst of phagocytic cells by nicotinamide adenine dinucleotide phosphate (NADPH) oxidase. This enzyme complex uses electrons derived from intracellular NADPH to generate superoxide anion, which is further processed to form hydrogen peroxide and other ROS-providing host defense against bacterial and fungal pathogens [ 2 ]. The essential role of the phagocytic oxidase in host defense is well illustrated by the serious phenotype of chronic granulomatous disease (CGD), in which susceptibility to infections develops in the absence of a functional phagocytic oxidase [ 3 ].
Non-mitochondrial production of ROS was detected in various cell types, showing that intentional generation of ROS is a general feature of many tissues, rather than a unique function of phagocytes. Homology searches in human genome databases resulted in the discovery of six novel NADPH oxidase enzymes: Nox1, Nox3, Nox4, Nox5, Duox1 and Duox2 [ 4–6 ]. They all have at least partially similar structure and generate ROS in response to various stimuli. The physiological function of these proteins and their potential role in the pathogenesis of human diseases is currently under intensive investigation.
Structural aspects
The phagocytic oxidase is a multi-protein enzyme complex, consisting of a membrane-bound flavocytochrome and several cytosolic regulators [ 2 ]. The flavocytochrome b 558 has two parts: the Nox family member and ‘prototype’ gp91 phox (now designated Nox2) and the smaller subunit p22 phox . The N -terminal part of Nox2 is composed of six transmembrane segments. This region contains four conserved histidine residues, which are responsible for haem binding. The two haems, embedded within the channel of the six α-helices, enable the transfer of electrons through the membrane. The carboxyl terminus of Nox2 has motifs for binding the coenzymes NADPH and flavin adenine dinucleotide (FAD).
Nox1, 3 and 4 are similar in size and domain structure to gp91 phox [ 7–9 ]. Among them, Nox1 and Nox3 show the closest homology to Nox2, while Nox4 is more distantly related to Nox2. Nox5 is larger than Nox2 because it has four additional cytosolic EF hands (motifs from the parvalbumin E & F helices) on its N -terminus [ 10 ]. These motifs are responsible for the regulation of proteins by Ca 2+ . Duox1 and 2 are the largest members of the Nox/Duox family [ 11 ]. They are called Dual oxidases because they possess an N -terminal peroxidase-homology domain in addition to their C-terminal NADPH oxidase domain. Between the NADPH oxidase portion and the peroxidase-like domain there is an additional trans-membrane domain and two EF-hand motifs, representing targets of regulation by calcium ions.
Regulation
In phagocytic cells, in addition to flavocytochrome b 558 , which consist of gp91 phox and p22 phox , there are other regulatory components needed for ROS production: p47 phox , p67 phox , p40 phox and the small guanosine triphosphate (GTP) Rac2 [ 2 ] ( Figure 1 ). P47phox, p67phox and Rac2 are essential regulators of Nox2, since a deficiency in any of these proteins results in immunodeficiency with absent or diminished superoxide production [ 3 ] Unstimulated phagocytes have inactive cytochrome b 558 stored within intracellular membranes; during activation, the cytosolic regulators are translocated to the catalytic core. P67phox and Rac seem to regulate catalysis directly, while p47 phox is a classical adapter protein, linking p67 phox to the gp91 phox [ 2 ].
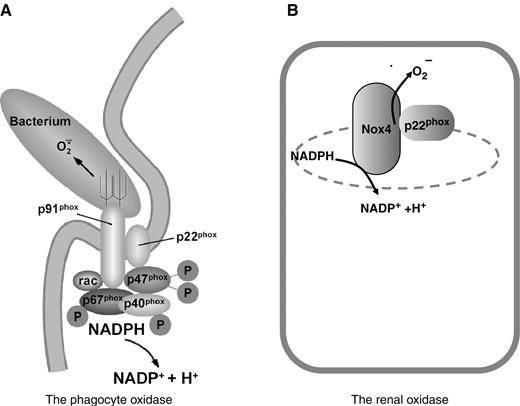
Molecular composition of the phagocyte- and renal-oxidase. ( A ) The phagocyte oxidase is a multicomponent enzyme which is localized to the phagosomal membrane during phagocytosis. ( B ) The renal oxidase contains Nox4 and probably p22 phox . The exact intracellular localization of Nox4 is currently unknown.
Our understanding of how other Nox/Duox proteins are regulated is far from complete, although recently it became evident that striking similarities exist between the regulation of phagocytic and non-phagocytic oxidases. First p22 phox , which is an essential partner of Nox2, can also form a functional complex with Nox1, Nox3, and Nox4 [ 12–15 ]. This finding explains previous studies, where high-level p22 phox expression was described in non-phagocytic tissues [ 16 ]. Nox5 and Duox proteins probably function independently of p22 phox , since Duox proteins are present and active in lower species where p22 phox homologues are not expressed (Geiszt et al ., unpublished data).
Based on the high structural homology between Nox1 and Nox2, it seemed likely that Nox1 is also regulated by cytosolic proteins. Indeed, the cytosolic components of the phagocytic NADPH oxidase (p47 phox and p67 phox ) could also increase the enzymatic activity of Nox1 [ 17 ]. Based on homology with p47 phox and p67 phox two novel regulator proteins were identified in colon epithelial cells, which are now designated as NOXO1 and NOXA1 [ 12 , 18 , 19 ]. NOXO1 is homologous to p47 phox and NOXA1 is homologous to p67 phox . Their function in the regulation of Nox1 was confirmed in heterologous co-expression experiments. The small GTPase Rac1 also regulates Nox1, since Noxa1 binds Rac1 in a GTP-dependent manner [ 12 ] and Nox1-mediated superoxide production is effectively reduced by mutant Rac1 proteins and Rac1-specific siRNA [ 15 , 20 , 21 ]. The activity of Nox1 is probably also regulated at the transcriptional level. In mesangial cells, nitric oxide (NO) inhibits the expression of Nox1, suggesting a crosstalk between NO and superoxide-generating systems [ 22 ].
The activity of Nox3 is also subject to regulation by cytosolic proteins. Data obtained from experiments with transfected cell lines suggest that Nox3 is able to produce low amounts of superoxide in the absence of cytosolic regulators [ 14 ]; however, its enzymatic activity is effectively stimulated by p47 phox and p67 phox [ 14 , 23 , 24 ]. NOXO1 is able to increase ROS production in Nox3-expressing cells, and this stimulatory effect seems to depend on p22 phox [ 16 , 24 , 25 ]. Recent genetic evidence supports the role of NOXO1 in the regulation of Nox3 [ 26 ]. Similarly to its role in the regulation of Nox1 and Nox2, Rac1 also regulates the activity of Nox3 [ 15 ].
We have little information on the regulation of Nox4. Nox4 seems to be constitutively active even in the absence of the cytosolic regulators, and expression of cytosolic Nox regulators in various combinations had no effect on the ROS output of Nox4-expressing cells [ 27 ]. As mentioned above, p22 phox can form a complex with Nox4, but it does not appear to serve as a docking point for the known cytosolic regulators [ 25 ] ( Figure 1 ).
Nox5 and Duox proteins all contain calcium-binding EF-hand motifs, suggesting that calcium signals that can arise in cells through several mechanisms regulate the activity of these enzymes. In fact, Nox5-expressing cells produce ROS when stimulated by calcium ionophores [ 10 , 28 ] and calcium is also an effective stimulus of different Duox-expressing cells [ 29 , 30 ]. Both Nox5 and Duox2 can be activated in cell-free systems by calcium [ 28 , 30 ]; in the case of Nox5, this appears to involve a direct intramolecular interaction of the calcium-binding EF-hands with the C-terminal domain [ 28 ]. It is possible that Nox5 and Duox proteins also have cytosolic regulators, but the identity of such proteins remains unknown. Grasberger and Refetoff [ 31 ] have recently identified Duox maturation factors (DUOXA1 and DUOXA2) which are necessary for the expression of functional Duox proteins.
Expression pattern
A common feature of the Nox/Duox family of oxidases is their widespread expression pattern, although they frequently exhibit their highest expression within specific tissues and cell types. Nox1 is highly expressed in colon epithelial cells, where it is readily detected by Northern blotting, in situ hybridization, and immunohistochemistry [ 7 , 17 , 32 ]. Besides its primary site of expression, Nox1 was also detected in several other cell types, including gastric mucosal cells [ 33 ], smooth muscle cells [ 7 ] and osteoclasts [ 34 ] The primary site of Nox3 expression is in the inner ear [ 23 , 35 ], where the enzyme is present in vestibular and cochlear epithelial cells and also in spiral ganglions [ 23 ]. The expression pattern of human Nox3 is currently unknown, but based on the high sequence homology between human and murine homologues, it is likely that in humans, Nox3 is also present in the inner ear. Lower level of Nox3 expression was detected in human fetal tissues, including fetal kidney [ 9 ]. Nox5 is expressed in lymphoid tissues such as spleen and lymph nodes and it is also present in the testis, where pachytene spermatocytes seem to express Nox5 [ 10 ]. Dual oxidases 1 and 2 were originally described in the thyroid gland [ 36 , 37 ], but subsequent studies have demonstrated that their expression is not limited to the thyroid, since they are present in many other tissues including the airways, salivary glands and gastrointestinal system [ 38–40 ]. In all of these tissues epithelial cells are the primary sites of Duox expression. The expression pattern of Nox4 will be detailed in a separate chapter.
Functional aspects
While Nox2 has a key role in antimicrobial host defense, the function of other Nox/Duox isoforms is far less understood. Many observations indicate that Nox1 could also have a role in host defense. The colon, where Nox1 is highly expressed, is inhabited by a variety of microorganisms and must therefore maintain an effective epithelial barrier against invasion of the host. In guinea pig gastric cells, Helicobacter pylori lipopolysacharides (LPS) induce elevated Nox1 and thereby produce higher ROS levels through toll-like receptor 4 (TLR) pathways [ 33 ]. Although colon epithelial cells lack this receptor, and do not react to LPS, Nox1-mediated ROS production in response to flagellin from Salmonella enteritides is mediated through TLR5 in these cells [ 33 ]. Viral introduction of Nox1 into Nox2-lacking CGD neutrophils can partly replace Nox2, and thereby partially rescue the deficiency in superoxide production, suggesting a role in innate immunity [ 17 ]. Nox1 expression in different colon epithelial cell lines is effectively up-regulated by the inflammatory cytokine IFN-gamma [ 17 , 41 ], which further supports a role for Nox1 in mucosal host defense.
Originally Nox1 was described as a ROS source that stimulates mitogenesis when over-expressed in NIH 3T3 cells [ 7 ]. Experiments performed on other cell lines did not support this early claim and found no effect of elevated Nox1 expression on mitogenesis [ 17 ]. Furthermore, studies on Nox1 expression in colon cancer samples found no positive correlation between Nox1 expression levels and proliferation or malignancy [ 17 , 42 , 43 ]; in two studies the more differentiated colon tumours were found to express higher Nox1 levels [ 17 , 42 ]. On the other hand, Lim et al . [ 44 ] found increased Nox1 expression and hydrogen peroxide production in prostate cancer samples.
Another intensively studied area of Nox1 function is its role in the vascular smooth muscle cells. Many agonists such as platelet-derived growth factor (PDGF), angiotensin II, prostaglandin F2α up-regulate Nox1 expression, whereas suppression of Nox1 by antisense techniques inhibits superoxide production in cultured smooth muscle cells [ 7 ]. Overproduction of ROS by Nox1 could lead to cell proliferation, hypertrophy, and, partly through degradation of NO, to increased blood pressure. Recently, three studies using transgenic animals suggested that Nox1 might be involved in the development of angiotensin II-induced hypertension [ 45–47 ]. Since Nox1 is also expressed in the kidney [ 22 , 48 ] future studies should focus on the potential role of renal Nox1 in the development of angiotensin II-induced hypertension.
Nox3 was recently shown to have a role in otoconia formation in the utricle and saccule of the inner ear [ 35 ]. Nox3-deficient mice are characterized by impaired balance and the lack of otoconia in the inner ear. The exclusive expression in inner ear (sensory epithelia and ganglia) corresponds with this observation [ 23 ].
Nox5 mRNA is expressed in testis, in pachytene spermatocytes, and in spleen, where it localizes to the B-cell rich mantle zone, surrounding germinal centers, and to periarterial lymphoid sheets [ 10 ]. Interestingly, lymphocytes from circulating blood are Nox5 negative. In lymphoid tissues, the physiological role of this calcium-regulated ROS production is unclear. Since host defense can be excluded, function in redox regulation of signalling pathways is the most likely explanation. In spermatocytes, a function in oxidative changes associated with sperm capacitation or the acrosomal reaction is suggested.
Duox1 and 2 were cloned from thyroid glands and were named at first as thyroid oxidases (thOX or tox) [ 36 , 37 ]. In the thyroid gland, the enzymes are used in iodide organification during thyroid hormone biosynthesis. The significance of two highly homologous Duox enzymes in the thyroid gland is not yet known. Patients with Duox2 deficiency have congenital hypothyroidism, which illustrates the essential role of Duox2 in thyroxine synthesis [ 49 ]. Duox enzymes are also present in other locations in the human body, where a role in hormone biosynthesis is not likely. It was proposed that in these tissues, Duox proteins would represent the missing, ROS-producing arm of the lactoperoxidase host defense system [ 38 ].
Nox 4, the renal oxidase
Nox 4 was first described as a renal-specific oxidase [ 8 , 50 ], and was thus originally named Renox. In situ hybridization experiments localized Nox4 mRNA expression to renal cortex in murine kidney, while the medulla showed much lower expression. Within the cortex, epithelial cells of proximal tubules express Nox4. Interestingly, in human kidney we detected Nox4 mRNA in medullary collecting ducts and also in epithelium on renal papillas ( Figure 2 ). Immunohistochemical studies also showed Nox4 expression in distal portions of the human nephron [ 50 ]. Nox4 is also found in mesangial cells of rat kidney [ 51 ]. Nox4 is expressed in many other tissues and cells including fetal liver, vascular endothelial cells, murine osteoclasts [ 8 , 52–54 ]. When over-expressed in NIH-3T3 fibroblasts, Nox4 increased superoxide production and decreased cell proliferation and induced the phenotype of cellular senescence [ 8 ]. The physiological function of Nox4 is currently unknown. Nox4 may serve different roles depending on the tissue where it is expressed and its function as a ROS producer. The expression pattern of Nox4 in the kidney is consistent with several renal-specific functions. We originally suggested that Nox4 might be the long-sought oxygen sensor that could regulate erythropoietin (EPO) production in the kidney. This hypothesis is supported by several findings. The potassium channel TASK-1, which is expressed in oxygen-sensing cells, is inhibited by hypoxia and the oxygen sensitivity of TASK-1 seems to be modulated by the activity of Nox4 [ 55 ]. Nox2, the phagocytic homologue of Nox4, was described as an oxygen sensor in the lung [ 56 ]. In the murine kidney, EPO synthesis occurs in the proximal tubules [ 57 ] or in close proximity to proximal tubules [ 58 ], which would make Nox4 an ideally located sensor to regulate this process. Although proline hydroxylases are now considered the primary sensors of oxygen and regulators of hypoxia-inducible factor-1alpha (HIF) levels [ 59 ], recent observations suggest that Nox4 can also regulate HIF through ROS production [ 60 ]. Furthermore, there are transcription factors such as GATA-2 that are H 2 O 2 -sensitive targets, that regulate EPO expression in a HIF-independent manner [ 61 ]. Finally, it was shown in recent studies that superoxide dismutase 3, which is also present in the proximal tubules, has a role in the regulation of EPO production [ 62 ].
Tissue distribution, protein interactions and functions of Nox/Duox enzymes [ 4–6 ]
| Enzyme | Other names | Site of expression | Protein interactions | Function |
|---|---|---|---|---|
| Nox2 | gp91 phox | Myeloid cells | p22 phox , p47 phox ,p67 phox , Rac1, Rac2 | Host defense Signalling? |
| Nox1 | Mox1, NOH-L | Colon, VSMC, prostate, uterus | p22 phox , NOXO1, NOXA1, Rac1 | Host defense? Blood pressure regulation |
| Nox3 | Inner ear, fetal tissues | p22 phox , NOXO1 , Rac1 | Otoconia Biosynthesis | |
| Nox4 | Renox | Kidney, endothelium, osteoclast | p22 phox | Oxygen sensing?, Vasoregulation? |
| Nox5 | Lymph nodes, spleen, testis | Signalling? | ||
| Duox1& 2 | Thox, Tox, LNOX | Thyroid, lung, salivary glands, gastro-intestinal tract | Hormone synthesis Host defense? |
| Enzyme | Other names | Site of expression | Protein interactions | Function |
|---|---|---|---|---|
| Nox2 | gp91 phox | Myeloid cells | p22 phox , p47 phox ,p67 phox , Rac1, Rac2 | Host defense Signalling? |
| Nox1 | Mox1, NOH-L | Colon, VSMC, prostate, uterus | p22 phox , NOXO1, NOXA1, Rac1 | Host defense? Blood pressure regulation |
| Nox3 | Inner ear, fetal tissues | p22 phox , NOXO1 , Rac1 | Otoconia Biosynthesis | |
| Nox4 | Renox | Kidney, endothelium, osteoclast | p22 phox | Oxygen sensing?, Vasoregulation? |
| Nox5 | Lymph nodes, spleen, testis | Signalling? | ||
| Duox1& 2 | Thox, Tox, LNOX | Thyroid, lung, salivary glands, gastro-intestinal tract | Hormone synthesis Host defense? |
Bold types mark functions and protein interactions which are supported by genetic evidence.
Tissue distribution, protein interactions and functions of Nox/Duox enzymes [ 4–6 ]
| Enzyme | Other names | Site of expression | Protein interactions | Function |
|---|---|---|---|---|
| Nox2 | gp91 phox | Myeloid cells | p22 phox , p47 phox ,p67 phox , Rac1, Rac2 | Host defense Signalling? |
| Nox1 | Mox1, NOH-L | Colon, VSMC, prostate, uterus | p22 phox , NOXO1, NOXA1, Rac1 | Host defense? Blood pressure regulation |
| Nox3 | Inner ear, fetal tissues | p22 phox , NOXO1 , Rac1 | Otoconia Biosynthesis | |
| Nox4 | Renox | Kidney, endothelium, osteoclast | p22 phox | Oxygen sensing?, Vasoregulation? |
| Nox5 | Lymph nodes, spleen, testis | Signalling? | ||
| Duox1& 2 | Thox, Tox, LNOX | Thyroid, lung, salivary glands, gastro-intestinal tract | Hormone synthesis Host defense? |
| Enzyme | Other names | Site of expression | Protein interactions | Function |
|---|---|---|---|---|
| Nox2 | gp91 phox | Myeloid cells | p22 phox , p47 phox ,p67 phox , Rac1, Rac2 | Host defense Signalling? |
| Nox1 | Mox1, NOH-L | Colon, VSMC, prostate, uterus | p22 phox , NOXO1, NOXA1, Rac1 | Host defense? Blood pressure regulation |
| Nox3 | Inner ear, fetal tissues | p22 phox , NOXO1 , Rac1 | Otoconia Biosynthesis | |
| Nox4 | Renox | Kidney, endothelium, osteoclast | p22 phox | Oxygen sensing?, Vasoregulation? |
| Nox5 | Lymph nodes, spleen, testis | Signalling? | ||
| Duox1& 2 | Thox, Tox, LNOX | Thyroid, lung, salivary glands, gastro-intestinal tract | Hormone synthesis Host defense? |
Bold types mark functions and protein interactions which are supported by genetic evidence.
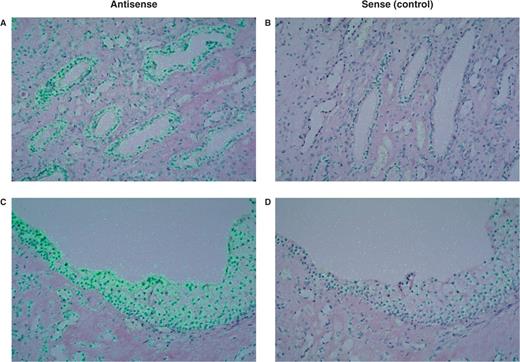
Detection of Nox4 mRNA in the human kidney medulla by in situ hybridization. Antisense ( A , C ) and sense ( B , D ) probes demonstrate specific expression of Nox4 mRNA in cells of medullary collecting ducts ( A ) and in epithelial cells of renal papilla ( C ). Superimposed polarized epi-illumination and bright-field images (in which the hybridization signal appears green).
It is also possible that Nox4, similar to its phagocytic counterpart, has a role in host defense. The expression pattern detected in the human kidney ( Figure 2 ) would support such a function, since Nox4 is mainly expressed in the medullary part of the human kidney. Producing microbicidal ROS at that locus of the kidney would provide an effective defense against infectious pathogens, which usually originate from the lower urinary tract. In support of this idea, it has been known for years that the urine contains hydrogen peroxide in relatively high concentrations [ 63 ].
Nox4 might also have a role in the development of the renal complications of diabetes mellitus. Etoh et al . [ 64 ] showed increased expression of Nox4 and p22 phox (both mRNA and protein) in the kidney of streptozotocin-induced diabetic rats. The localization of the two proteins was in parallel with the accumulation of 8-hydroy-deoxyguanosine (8-OHdG), which is a marker for ROS-induced DNA damage. In the same animal model of diabetes, antisense oligonucleotides targeting Nox4 mRNA were effective in reducing the hypertrophy and fibronectin expression [ 65 ], which are characteristic markers of diabetic nephropathy. These data point to a signalling role for Nox4-derived ROS in the development of diabetic nephropathy, but the specific target(s) of ROS remain to be identified.
Nox4 was suggested to participate in insulin receptor signal transduction [ 66 ]. In many cells, growth factors and insulin stimulate low-level hydrogen peroxide production. Hydrogen peroxide then inhibits tyrosine phosphatases, thus enhancing the tyrosine phosphorylation induced by the receptor agonists [ 67 ]. Recently, it was suggested that Nox4 is responsible for insulin-induced H 2 O 2 production in 3T3-L1 adipocytes [ 66 ]. Gorin et al . [ 51 ] showed that Nox4 is involved in angiotensin II-induced ROS production in kidney mesangial cells. Since PDGF receptor stimulation also involves ROS production and enhanced cellular protein phosphotyrosine levels, a role for Nox4 in this receptor pathway was also suggested recently [ 68 ]; the decreased growth factor responses of fibroblasts from patients with Leprechaunism was attributed to down-regulation of Nox4.
Nox4 may have important roles in the cardiovascular system. According to expression studies performed on vascular endothelial cells, Nox4 seems to be the dominant ROS source in endothelial cells [ 53 , 69 ]. This localization is particularly exciting, because it would mean that Nox4-produced superoxide could effectively antagonize the effect of NO produced in the same cell type. A recent publication [ 53 ] has located endogenous Nox4 within the nucleus of human umbilical endothelial cells (HUVECs). The nuclear fraction of HUVEC cells produced superoxide in a NADPH-dependent way. Earlier reports already pointed to the nucleus as an intracellular site of ROS production in endothelial cells [ 70 ], and NADPH oxidase components were detected in nuclear fraction of endothelial cells [ 70 ] and B lymphocytes [ 71 ]. The nuclear localization of Nox4 suggests that it could convey redox signals altering gene expression. This suggestion is now supported by some experimental evidence [ 53 ], but the idea requires more rigorous testing. In blood vessels, Nox4 is also present in smooth muscle cells, where it localizes to the endoplasmic reticulum and nucleus [ 72 , 73 ]. 7-ketocholesterol, a major oxysterol component in low-density lipoprotein, stimulates the expression of Nox4 in smooth muscle cells, thus, ROS production by Nox4 might be responsible for the oxidative stress induced by 7-ketocholesterol [ 72 ].
Nox4 is also present in the heart, where cardiac fibroblasts express the enzyme. Transforming growth factor (TGF)-β stimulates the conversion of fibroblasts into myofibroblasts in a ROS-dependent manner. Nox4 is the likely source of oxidants in this process, since down-regulation of Nox4 expression by siRNA inhibited both ROS-production and the TGF-β induced expression of smooth muscle actin [ 74 ]. The involvement of Nox4 in TGF-β signalling was recently described in human pulmonary artery smooth muscle cells and also in HUVECs [ 75 , 76 ].
Nox4 is also expressed in cells of haematopoietic origin. CD34+ cells exhibit significant extra-mitochondrial oxygen consumption, which might be explained by the activity of Nox2 and Nox4, which are both detected by reverse transcriptase PCR [ 77 ]. The significance of Nox2 expression in haematopoietic stem cells is unclear, since no defect in blood cell formation was described in Nox2-deficient CGD patients or in animal models of CGD. Another extra-renal site of Nox4 expression is the murine osteoclast, where Nox2 is also detected [ 54 ]. It is possible that Nox4 and Nox2 together provide oxidative support for bone resorption, although this hypothesis requires further investigation.
Conclusion
With the discovery of novel NADPH oxidase isoforms, a very exciting area has began in ROS research. On the one hand, in tissues where the production of ROS has already been described (thyroid gland), we can now attribute this biological response to the activity of specific enzymes, allowing a more thorough understanding of the ROS-production process. On the other hand, the expression of these enzymes at sites where ROS-production has not been studied before expands our knowledge of the importance of ROS signalling. The exact function of the non-phagocytic Nox/Duox proteins remains to be explored; however, it now seems likely that different epithelial cells, particularly along mucosal surfaces, use these proteins—especially Nox1, Duox1 and 2—for ROS-based host defense. High-level ROS production at specific sites by non-phagocytic oxidases suggests that they have additional unique functions, such as hormone biosynthesis in thyrocytes (Duox2) or formation of otoconia (Nox3). The involvement in signalling is also an important question that must be studied further. Among new challenges are the collection of more data on the molecular assembly of these novel enzymes and the identification of their role in disease pathogenesis. It is likely that with broader knowledge we will be able to define the role of these novel ROS-sources in different disease processes, which can arise from either under- or over-production of ROS.
Acknowledgements
Experimental work in the authors’ laboratory was financially supported by grants from the Hungarian Research Fund (OTKA 042573) and the Cystic Fibrosis Foundation. M.G. is recipient of a Wellcome Trust International Senior Fellowship.
Conflict of interest statement . None declared.


{kind=link}
{kind=link}
Comments